| Новости | Конференция | Чат | База данных | Творчество | Сообщество | О сайте | English |

|
Помощь сайту |
|
ВВЕДЕНИЕ В КСЕНОБИОЛОГИЮ 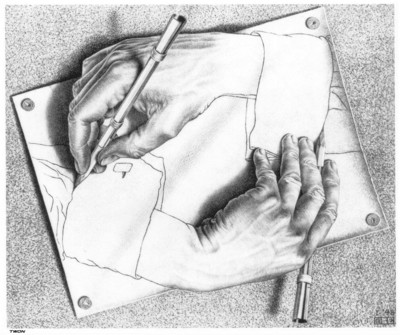 Когда наш мир на треть был меньше, На треть синей и горячей, Погасших звёзд алмазный пепел Расцвёл в тепле чужих лучей. Виктор Аргонов, техноопера «Русалочка» Вместо предисловия Теперь, когда с помощью орбитального телескопа Кеплер открыто уже более двух тысяч планет за пределами Солнечной системы, а в следующем десятилетии ожидается прорыв в изучении состава атмосфер наиболее близких из них при помощи строящихся наземных экстремально больших телескопов: прежде всего Гигантского Магелланового и орбитального телескопа им. Дж.Вебба, который должен придти на смену телескопу им. Хаббла, вновь обостряется вопрос о жизни за пределами Земли. Но, как и полвека назад, всякий, собирающийся написать книгу, сценарий или же разрабатывающий сюжет к игре в жанре строгой научной фантастики и при этом имеющий намерение описать «экзотические» формы жизни, т.е. организмы, фундаментально отличающиеся по своей химической архитектуре, либо вообще основанные на каких-то иных формах материи или даже самих принципах организации сложных систем, встает перед пугающей бездной почти полной неопределенности: ведь жизнь известна нам пока только в одной форме, а по одной точке адекватный график построить невозможно. В то же время хорошо известные общие свойства нашей Вселенной, уже давно открытые как естественными термодинамикой, ядерной физикой и химией, так и «сверхъестественной» математикой, да и просто ключевые закономерности развития известной нам жизни, налагают очень серьезные ограничения на многие наши фантазии. И по этим ограничениям теперь вполне можно хотя бы в самых общих чертах построить коридор «поканезапрещенных вариантов». Тогда на него могли бы ориентироваться те, кто в своем творчестве не хотел совершенно отрываться от физической реальности. Этим академически-честным творцам прежде всего и предназначается наша работа. Если составить перечень самых фундаментальных вопросов, стоящих перед человеческой цивилизацией, то проблема происхождения и единственности или, наоборот, неединственности жизни в нашей вселенной всегда будет в числе первых трёх, которые обозначил в заглавии своей известнейшей книги ещё И.Шкловский — «Вселенная, жизнь, разум». Значение того, каким будет ответ на вопрос «Одиноки ли мы во вселенной?» в истории цивилизации исчерпывающе передано, например, А. и Б. Стругацкими в самом начале «Сталкера»: "Сам факт Посещения является наиболее важным открытием не только за истекшие тринадцать лет, но и за все время существования человечества. Не так уж важно, кто были эти пришельцы. Неважно, откуда они прибыли, зачем прибыли, почему так недолго пробыли и куда девались потом. Важно то, что теперь человечество твердо знает: оно не одиноко во Вселенной. Боюсь, что институту внеземных культур уже никогда больше не повезет сделать более фундаментальное открытие." Разумеется, обнаружение жизни разумной (о которой идет речь и в приведенной цитате) будет иметь ничуть не меньшее значение, но какой вопрос бы мы ни поставили: возможен ли (и насколько) во вселенной иной разум или иная жизнь, сразу за ним встает другой: а что именно мы понимаем под этими словами? При ближайшем рассмотрении практически любого природного явления оно неизбежно распадается на более простые элементы и нам придется решать: какие из них мы должны считать ключевыми? Рассматриваемых явлений, как наиболее сложных из известных нам, это касается в наибольшей степени. Особенно плохо обстоит дело с «разумом», где разнообразие «ключевых свойств» простирается от способностей к решению логических задач и созданию орудий до уровня социальных взаимодействий и языка. А по мере того, как стараниями этологов расширились наши знания о мыслительных способностях братьев наших меньших, ближайших и не только, круг свойств этих изрядно сузился, а сами они стали во многом количественными и сейчас из объективных поводов для «видовой гордости» навскидку можно вспомнить разве что число регистров рабочей памяти более четырёх, что в сочетании с высокими социальными способностями дало число Данбара более нескольких десятков, да язык со сложной грамматикой. Поскольку интеллект в широком ссысле — как способность к опережающей регуляции в виде увеличения числа степеней свободы для себя в будущем (что ещё С.Лем в Сумме Технологии назвал «гомеостатом второго рода») и выраженной теперь математически подобно уравнению энтропии Больцмана Висснер-Гроссом можно наблюдать даже на примере эволюции генофонда популяции бактерий (что, кстати, тоже было предсказано С.Лемом в псевдопредисловии Эрунтика сборника «Мнимая Величина»). Задача же определения критериев «жизни вообще», о которой только и пойдет речь далее, кажется куда более простой. В целом это, конечно, так и есть, но распада нашего интуитивного понятия при его формализации на составляющие избежать не удается и здесь. Более того: при погружении в биологию уже чуть глубже среднешкольного уровня становится ясно — насколько разным по уровню сложности бывает то, что мы можем называть живым. Кроме того: даже однажды появившись, вовсе не факт, что жизнь будет способна эволюционировать до той сложной формы (эукариотических клеток), что мы повсюду наблюдаем на нашей планете половину её истории (около двух миллиардов лет). И даже если запустить её заново в тех же начальных условиях — также совсем не обязательно, что возникнут и будут развиваться именно те же линии многоклеточных растений и животных, которые известны в последние полмиллиарда лет. Таким образом, надо всегда помнить, что вполне возможна такая ситуация: неспециалист, узнав подробности об однажды и даже неоднократно открытых, но очень просто устроенных внеземных организмах, может воскликнуть: «Да разве-ж это жизнь?!»,- несмотря на то, что по всем нашим фундаментальным критериям мы должны будем признать их живыми. Со времен наиболее известных первопроходцев тут — Шредингера и Шкловского, Опарина и Холдейна — утекло немало жидкостей, и в этой части картины мира многое обрело существенно иной вид. Поэтому нам придется посмотреть на то, как феномен жизни выглядит теперь в свете новейших достижений самых разных отраслей естественных наук. Хотя проблема абиогенеза (происхождения жизни) в настоящее время в мире изучается очень активно и продуктивно, область допускаемых современной академической парадигмой вероятностей и вариантов жизни по сей день остается чрезвычайно широкой. Она простирается от «панбиологического ультраоптимизма» (обычно свойственного небиологам и нехимикам, начиная с Артура Кларка и его термина водно-углеродный шовинизм) в выпущенном семь лет назад 100-страничном докладе Национальной академии наук США «Пределы органической жизни в планетарных системах» до «тотального антропного скептицизма» в вышедшей три года назад книги ведущего исследователя Национального Центра Биотехнологической Информации Евгения Кунина «Логика Случая» , в прошлом году переведенной и дополненной автором для русскоязычного читателя. Более популярных, но столь же широких, подробных и современных работ по этой теме на русском языке исчезающе мало: это главным образом популярные статьи Александра Маркова и Михаила Никитина, а также изданная в 2000 году и переиздающаяся поныне книга Кирилла Еськова История Земли и жизни на ней, на которых по большей части и основана эта работа. 0. Математика биологии (логические ограничения) Итак, если мы собираемся заниматься ксенобиологией, т.е. искать жизнь-какой-бы-она-ни-была, то нам прежде всего нужно определить — что же собственно такое в нашем понимании «жизнь». Причём, по возможности, максимально формализованно. Уже традиционно любое погружения в аксиоматику биологии начинается с известнейшего высказывания одного из основателей синтетической теории эволюции — Феодочия Добжанского: «Ничто в биологии не имеет смысла кроме как в свете эволюции». Причём эволюции в основе своей дарвиновской. Что это значит? Это значит дифференциальное выживание наиболее приспособленных (к конкретным условиям) вариантов. Совершенно очевидно, что количество информации в живой системе должно быть больше чем один или даже несколько бит — только в этом случае она может стать объектом изучения информатики или биологии, а не только физики или химии. Раздел между этими науками лежит именно в комбинаторной сложности исследуемых ими явлений: в ряду теорфизики, физики элементарных частиц, ядерной физики, неорганической химии, органической химии, биохимии, микробиологии, ботаники и зоологии по меткому выражению Э.Резерфорда уменьшается количество собственно «физики»(разнообразие фундаментальных взаимодействий) и увеличивается «коллекционирование марок»(число возможных сочетаний элементов сходной природы), которое уже в случае гетероорганических (т.е. содержащих любые другие элементы кроме углерода и водорода) соединений и гетерополимеров на многие порядки превышает число элементарных частиц в наблюдаемой Вселенной (и поэтому не может быть перебрано за все время ее существования). Поскольку любые системы хранения информации, которые способна разработать природа(как минимум) в нашей Вселенной никак не могут обеспечивать пассивное хранение информации дольше двух-трех сотен миллионов лет, то информация, претендующая сохраняться дольший срок, обязана прибегать к резервному копированию. Хотя активная работа по поддержанию сложной структуры требует потребления свободной энергии из среды, «пассивные хранители» в этой среде будут весьма доступным источником и энергии и стройматериалов. Отсюда сразу возникает конкуренция, ещё более усложняющая судьбу «хранителей», а если ещё и учесть их неспособность к эволюции, то очевидным становится факт, что в основе любых долгоиграющих хотя бы миллиард лет и хоть сколь-нибудь сложных структур, т.е. того что мы и зовем жизнью (а не кристаллами или иридиевыми DVD-дисками) всегда лежит процесс активного самокопирования. И только отчасти, живые системы прибегают к стратегии длительного пассивного сохранения, и то — только когда уже имеют весьма непросто устроенные и довольно энергетически затратные системы для этого (например — способность бактерий к спорообразованию). И даже в основе частичной регенерации тоже будет лежать всё тот же принцип самокопирования, но под контролем организма. И этот способ сам по себе таит риск (в случае многоклеточных — опухоли): «No system is safe» — принцип копирайта внутренне противоречив. Разумеется, что точность копирования должна быть достаточной для принципиально неограниченной серии размножения (т.е. живой организм должен быть полноценным автоматом фон Неймана), однако оно должно быть не совершенно точным — иначе при любом серьезном изменении условий среды (смене направления отбора) не будет ресурса для приспособления. Как следствие — система исходно должна иметь некий, причём не особо малый «запас свободы» вариантов себя. Эта граница между хаосом и порядком, между вырождением и бессмертием известна под названием порога Эйгена. Давно известен тот факт, что надежная передача информации возможна только на базе дискретных систем — неслучайно почти все современные информационные системы цифровые, хотя на заре вычислительной техники на аналоговые вычислители возлагались очень большие надежды. Поэтому и архитектуры живых организмов, которые способны першагивать порог Эйгена миллиарды лет, должны содержать информацию о своей структуре обязательно в дискретном виде (см. стр. 33-36). Справедливости ради следует упомянуть известный (и все еще спорный) казус с эволюцией прионов. Однако и количество пространственных конформаций белков — также конечно и дискретно, а в случае автокаталитической «репликации» прионов — весьма ограничено в диапазоне вариантов и возможно только на базе уже существующих живых систем, которые эти белки производят — этот пример наглядно демонстрирует «потенциал» аналоговой архитектуры в качестве основы для жизни и тем, кого не убеждает вышеприведенный строгий математический обзор Сергея Вакуленко: как говорится, sapienti sat. Что из этого следует для ксенобиологии? А вот что: если мы абстрагируемся от материальной природы живой системы и от того, будет ли она проявлять свою активность когда мы её изучаем, то мы должны искать любые высоконерегулярные последовательности (массивы) конечного числа элементарных блоков, но встречающиеся хотя бы в нескольких копиях и вариантах. Топология этого массива будет скорее всего одномерной — как классическая Машина Тьюринга и подобно нашим белкам и нуклеиновым кислотам; в самом оптимистическом варианте — двухмерной (как QR-код), поскольку только в этих случаях возможно его считывание путём непосредственного пространственного контакта. Тогда как информация из сплошного объемного блока может быть считана только при использовании алгоритмов томографии, нередуцируемая сложность материального воплощения которых (нужен сразу уже готовый и очень сложный вычислитель) не оставляет никакого реального шанса на самовозникновение в наблюдаемой Вселенной, не говоря уже о том, что неразрушающее считывание такого массива надмолекулярных масштабов может быть осуществлено только с помощью источника хорошо сфокусированного света (если объем прозрачен) либо ядерного магнитного резонанса. Ни то ни другое химическим репликаторам недоступно прежде всего ввиду энергетических и размерных ограничений. Таким образом совершенно однозначно получается, что настоящие живые организмы могут существовать только в виде конечных автоматов и, похоже, только Тьюринг-полных. И размножающихся по принципу матричного самокопирования. 1. Физика биологии (энергетические ограничения) Какие свойства характерны для живых систем с точки зрения физики — известно уже давно, начиная со Шредингера и заканчивая Пригожиным. Это прежде всего способность поддерживать термодинамическое НЕравновесие (метастабильность, гомеостаз) с окружающей средой, активно выкачивая из неё свободную энергию, ускоряя её энтропийное рассеивание (диссипацию). Согласно пригожинской неравновесной термодинамике , такие системы могут (но должны ли?) появляться путем самоорганизации только там, где есть достаточный (очень большой) запас свободной энергии (градиент). И с этой точки зрения первое, на что обращается внимание — это не планеты, а звезды. Но если посчитать удельное энерговыделение звезды, способной существовать хотя бы несколько миллиардов лет, то оно оказывается на порядок меньше удельного тепловыделения живого бодрствующего млекопитающего. А если ещё учесть энергию на синтез и распределение элементов с III периода, затраченную значительно более крупными звездами предыдущих поколений (а за железом — и сверхновыми!), то соотношение градиентов этих оказывается вовсе не в пользу звездной среды. Тем не менее физиками рассматривались вопросы самоорганизации плазменно-пылевых сред в метастабильные структуры, но только с помощью теоретического моделирования, и при этом не был показан вышеуказанный ключевой для биологии процесс — копирования массива информации. В любом случае при поиске жизни неизвестной природы следует придерживаться общего подхода, прямо следующего из положений предыдущей части: если будут обнаружены нерегулярные последовательности элементов/состояний в метастабильных структурах и при этом они будут слишком маловероятными для одномоментного самовозникновения, но будут выявлены хотя бы в нескольких копиях, причем более одной точной разновидности — то только в таких системах можно подозревать жизнь. Что же касается конкретных значений физических параметров, в которых может существовать активная жизнь, то для современной земной биоты диапазон давлений при невысоких температурах составляет 6 порядков, почти совпадая с диапазоном существования жидкой воды: от 2,5 кПа (вода при таком давлении кипит при 21°C) (однако давление на современном Марсе в три раза меньше и вода там кипит уже при 5°C); и за 2 ГПа — т.е. фактически вплоть до образования экзотического льда-VI с сохранением жизнеспособности в нём. А вот диапазон температур куда более ограничен (что неудивительно) и для активной жизни составляет 140 градусов: от -18°C (255K) в соленой воде, до 122°C (395K) в воде под давлением в 2,1 атм. Однако некоторые белки, а также ДНК в сухом виде или в виде сухих спор выдерживают до ~150°C. Середина этого диапазона приходится на 50-60°C и, судя по свойствам наиболее древних ферментов и температур плавления водородных связей самих нуклеиновых кислот, вполне вероятно, что наша жизнь возникла как раз при этих условиях. Расширить же этот диапазон получится не более чем в два раза: прибавив ещё по 70-75 градусов с каждой стороны, мы получим 175K (~-100°C) — температуру замерзания водно-аммиачной эвтектической смеси и большинства самых холодостойких полярных органических растворителей (почему именно их — см. конец главы «Химия биологии»), кроме этанола и н-пропанола; и 470K (~200°C) — предел термостойкости для большинства полимеров-пластиков, кроме ароматических полиамидов и некоторых силиконов. И не следует думать, что эти «взятые с потолка» пределы можно превзойти, поскольку даже они — уже крайне оптимистическая оценка, по причине того, что в основе жизни лежат сложные постоянные структуры, для которых условия существования — заведомо уже. Ровно то же самое, кстати, касается и чаемых многими футурологами молекулярных машин-наноассемблеров, работа которых по принципу так называемого механосинтеза (если он вообще пригоден для получения активных механизмов) скорее всего будет абсолютно ограничена сверху, а подобного живым клеткам «синтеза в растворе» — снизу, температурой в районе 145K (-128°C) в области «вменяемых» давлений (до 10 ГПа), за девять градусов до которой сегодня заканчивается мир чистых квантовых явлений в виде рекорда выскотемпературной сверхпроводимости (138К) и начинается мир тотального броуновского движения в виде точки плавления самого холодостойкого полярного растворителя — н-пропанола (147K). Верхний же предел всякой активной сложности на молекулярном уровне обозначает температура, при которой начинают распадаться ароматические полиамиды и силиконы — порядка 570K (~300°C) — достижимая уже на хорошей электрической или газовой плитке. Хотя очень многие простые организмы очень хорошо сохраняются в замороженном или высушенном виде не теряя жизнеспособности, тем не менее, сохранение их дольше четверти миллиарда лет вряд ли возможно — прежде всего в результате воздействия практически неустранимой в природных условиях фоновой радиации, в отношении которой рекорд выносливости современной активной земной жизни составляет 30 тысяч Грей для простых организмов и 6 тысяч Грей для сложных. С другой стороны, жизнь, не смотря на некоторую ограниченность доступных ей элементов и отставание на многие порядки по отдельным показателям от больших и узкоспециализированных агрегатов, порожденных цивилизацией, добилась очень впечатляющих успехов в области эффективности манипуляций на молекулярном уровне, вплотную подойдя к термодинамическому пределу возможного: так, делящаяся каждые ~20 минут кишечная палочка всего в 6 раз менее эффективна, чем это возможно теоретически, причем существуют и более быстрые бактерии, рекорд скорости деления которых в свежей оптимальной среде составил чуть менее 7 минут. Кроме того, известно как минимум восемь широко распространенных каталитически-совершенных ферментов (в основном занимающихся «выключением» опаснейших форм кислорода, распространенного антибиотика и основного нейромедиатора), каждый пространственный контакт которых с молекулой исходного вещества катализирует соответствующую реакцию. А менее совершенные ферменты стало получаться превосходить только совсем недавно, причем, что характерно — при помощи одного из самых редких и трудных в обработке элементов — иридия. С учетом только вышеизложенных фактов, очень скептичным становится взгляд не только на попытки найти более совершенные формы жизни в природе, но и на опасение превзойти её на порядки с помощью наноинженерии, известное под термином «серая слизь». По всей видимости, превзойти известные живые организмы сразу по множеству параметров и с помощью столь же универсальных молекулярных автоматов если и удастся, то только в те же полтора-два раза, но никак не на порядок. Постоянная Больцмана и энергия химических связей — это не те вещи, которыми можно пренебрегать как при построении моделей возникновения и развития жизни, так и для предсказания возможностей любой машинерии: хоть молекулярного, хоть астрономического масштаба. 2. Химия биологии (комбинаторные ограничения) Таким образом, вся известная нам физика ограничивает существование достаточно сложных для возникновения жизни структур только областью низких энергий, где существуют химические соединения. И прежде всего нас будет интересовать аналог самокопирования в химии — автокаталитические (цепные) реакции. Реакции эти довольно распространены и аналоги их можно подобрать и в ядерной физике, но к жизни они не имеют отношения до тех пор, пока при автокатализе не происходит образование структур такой сложности, одномоментное самовозникновение которых в данной среде исчезающе маловероятно (т.е. содержать массив информации). Но следует заметить, что возникновение жизни вообще все таки может быть не столь уж пессимистично-маловероятным в принципе, особенно в свете того, что эволюционный потенциал переходного — предбиологического — этапа нам пока еще очень мало понятен. Тут вполне могут быть полезными, например, давно известные свойства палиндромных последовательностей. Что же касается альтернативной биохимии, то тут, несмотря на видимое разнообразие таблицы химических элементов Д.И.Менделеева, прочие описанные там «экзотические» комбинации либо невозможны в чистом виде, либо уже и так задействованы в известной нам жизни. И даже в случае возникновения такой «экзотики» скорее всего их эволюционный потенциал и вероятность возникновения существенно меньше, чем у нашей. А накладываемые химией ограничения и минимальные прямые аналогии позволяют нам представить хотя бы общие очертания этих возможностей. Почти все теоретики согласны с тем, что главных химических «ингридиентов» жизни должно быть как минимум два сорта: - те, из которых строятся сами биомолекулы - и те, которые образуют среду для этого строительства. Первые должны быть способны образовывать большие, сложные и в то же время достаточно стойкие молекулы, а вторые — обеспечивать их хорошее перемешивание (диффузию) в равномерных условиях, что возможно только в жидкостях. Кристаллические формы жизни невозможны по очень простой причине: без движения невозможно физическое самокопирование, а запись в виде состояний ячеек памяти (как в чипах) никогда не выйдет за пределы доступной памяти (как те же прионы из первой главы) и не сможет предотвратить их разрушение. Механическое же движение исходно очень сложно и затратно по сравнению со статистическими, но «дармовыми» температурными броуновскими колебаниями в жидкости. С другой стороны газовая среда обеспечивает слишком малую вероятность химических взаимодействий и не способна удерживать «на плаву» сколь-нибудь крупные структуры. Итак, что касается биомолекул: сложные, нерегулярные и в то же время высокостабильные структуры получаются только с помощью элементов, способных связываться между собой прочными ковалентными связями в длинные ветвящиеся цепочки. На эту роль подходят только неметаллы с валентностью больше двух (для ветвления цепей): бор, углерод, азот, кремний и фосфор. Но бора при термоядерном синтезе в естественных условиях образуется очень мало, азот способен к полимеризации только при сверхвысоких давлениях глубоко в недрах планет-гигантов, до которых вряд ли когда-либо удастся добраться, а элементы третьего периода — кремний и фосфор — формируют стабильные цепи только при чередовании с кислородом, причём гораздо более массивные и склонные к образованию однообразных сетчатых (двухмерных) полимеров. Кроме того, эти элементы в цепи не обладают способностью образовывать кратные пи-связи, совершенно необходимые для сборки плоских структур, поэтому «чисто-силоксановая» жизнь невозможна — совсем без углерода обойтись не удастся и максимум возможного здесь — примесь кремния в гетероорганических молекулах: органосилоксанах и, возможно, органосилатранах. А фосфор и так уже запользован нашей жизнью для связи нуклеотидов в нуклеиновых кислотах и одновременно как оперативное хранилище химической энергии в виде свободных нуклеотдов АТФ и ГТФ. Более того: если мы обратимся к составляющим основу и нашей жизни — якобы «строго-углеродным» главным деталям — азотистым осонованиям нуклеиновых кислот и пептидам — то там мы увидим, что в их скелете (центральной цепи атомов) практически на каждые два атома углерода приходится один атом азота: Рис.1. Природные нуклеотиды ДНК (вид с торца) (A) и фрагмент пептидной цепи (B). Зеленый цвет — атомы углерода, синий — азота, красный — кислорода 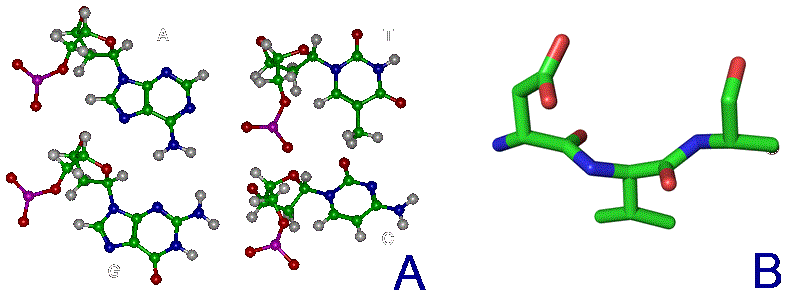 Так что с этой точки зрения куда правильнее было бы называть нашу жизнь «углерод-азотной». Кроме того не следует забывать про большое количество кислорода в углеводах и серу, хотя бы пара атомов которой есть практически в любой более-менее сложной белковой молекуле. Еще одно ограничение, накладываемое на кремниевую жизнь состоит в том, что синтезу кремний-органики довольно сильно препятствует наличие воды, а это в целом — гораздо более распространенное соединение во Вселенной, чем серная кислота и даже оксид кремния(песок и камень), не говоря уже о фторводороде — почти столь же редком, как и соединения бора. Все это находится в полном соответствии с приведенным выше «математическим» определением жизни — согласно ему в основе жизни должны лежать довольно стабильные и достаточно точно копирующиеся нерегулярные последовательности конечного числа блоков, а вот скелет цепи этих блоков должны составлять уже регулярные чередования разных атомов: хотя достаточно длинные однородные, «гомоатомные» цепи и могут существовать (пример — нонаконтатриктан), операции с ними (синтез, обратимое разделение и сшивка) «подручными средствами» (без широкого изменения физических параметров среды) крайне затруднены. Для упрощения понимания можно провести такую аналогию: наши методы химического синтеза работают подобно заводским пресс-формам или как минимум 3D-принтеру — довольно большому и сложному механизму, производящему монолитные вещи, тогда как биополимеры гораздо более напоминают LEGO-конструктор: если у вас есть только один тип блоков — вы многого сделать не сможете, а чтобы блоки можно было легко перекомбинировать, сами они должны иметь неоднородность, но повторяющуюся (паз-выступ). Регулярные цепи из разных элементов — это и есть полимеры. Аналогию с LEGO можно даже углубить: всякий, кто пытался собрать что-то действительно сложное из такого конструктора, знает как ценны в наборе детали, имеющие боковые выступы (так называемые SNOT-элементы). Совершенно очевидно каждому: как повышает разнообразие конструкций «свобода маневра» в трех измерениях. Элементарные детали обоих наших биополимеров — нуклеиновых кислот и белков — нуклеотиды и аминокислоты — устроены именно как SNOT-элементы LEGO, хотя их боковые цепи не позволяют ветвить скелет (впрочем, и в обычных конструкторах эти связи прочностью обычно не отличаются, чему в макромире виной гравитация): Рис.2. Элементарные звенья известных биополимеров. Слева направо: нуклеотид, аминокислота, SNOT-элемент. Стрелкой показано направление роста цепей. 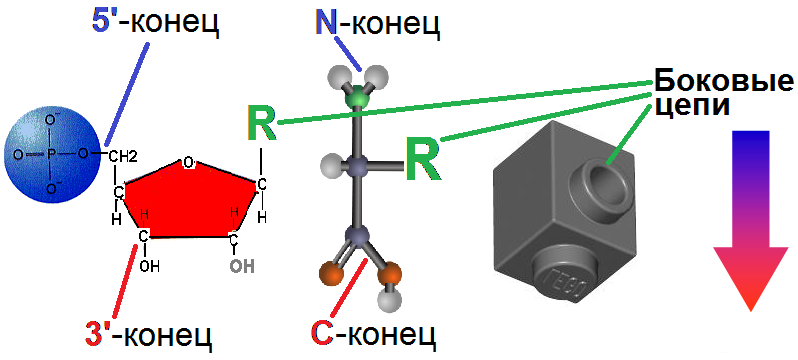 Отсюда же и произрастает явление зеркальной несимметричности (хиральности) биомолекул: боковые ветви симметричной цепи дают разнообразие только в двух измерениях. При этом наши биополимеры обладают тут интересной особенностью: нуклеиновые кислоты и белки живых организмов состоят только из мономеров одной из зеркально-симметричных форм, тогда как при обычном химическом синтезе обе формы образуется практически в равных соотношениях — это так называемая проблема хиральной чистоты биополимеров (homochirality). Биологическое значение этой избирательности очевидно: такая унификация сильно упрощает обеспечение пространственной (третичной) структуры этих макромолекул, от чего прямо зависит их функция. Кстати: так легко объясняется загадочная на первый взгляд спиральная упорядоченность многих биологических объектов начиная с ДНК: это просто многократно повторенная в одну и ту же сторону асимметричность составляющих её элементов, причём усложняющая ей жизнь (расплетение этой спирали при копировании (репликации) требует специальных ферментов — топоизомераз), поскольку никакой более сложной пространственной (третичной) структуры ДНК не образует, что ей, как пассивному хранилищу информации, и не требуется. Но требовалось её прямому предшественнику — РНК в эпоху «РНК-мира», когда один тип молекул нес на себе функции и хранения и реализации наследственности . РНК же третичную структуру очень даже имеет, активно формируя связи не только между двумя отдельными, полностью комплементарными цепями, но и в пределах одной цепи, также как и белки образуя сложные трехмерные «загогулины», что было бы невозможно, если бы составляющие её звенья-нуклеотиды были симметричными. Это наглядно видно даже в структуре самых маленьких цепочек — транспортных РНК наших клеток: Рис.3. Свернутая в трехмерном пространстве(третичная) и «расплющенная» на плоскости(вторичная) структуры молекулы транспортной РНК. 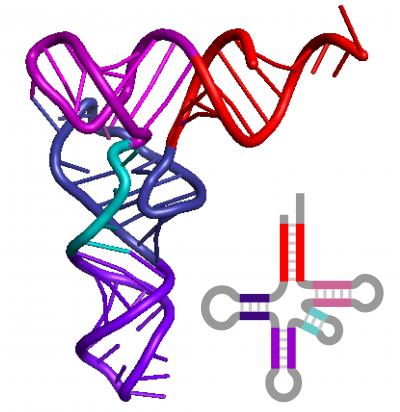 Таким образом «завораживающую спиральность ДНК» можно рассматривать просто как наследие (рудимент) РНК-мира. Механизмы «очистки» исходной смеси изомеров теперь понемногу проясняются, но первопричина и вероятность выбора жизнью той или иной формы — пока остаются не до конца ясными. Можно сказать, что тут существует три подхода: 0) «Нулевая гипотеза» — первая молекула РНК-репликатор должна иметь либо правую, либо левую хиральность, но произошло это чисто случайно (как подброшенная монета не упадет на ребро) — а дальше заработала биологическая эволюция по принципу «кто первый встал — того и тапки». 1) Влияние космического циркулярно-поляризованого ЭМ-излучения. 2) Гипотеза Вестера–Ульбрихта, согласно которой гомохиральность жизни является следствием фундаментальной пространственной несимметричности слабого взаимодействия элементарных частиц — значительно менее популярная среди специалистов версия. Кроме того, не так давно была опубликована работа, которая хотя и не пытается решить «в лоб», а очень изящно ставит эту «проблему» не в начало абиогенеза, а в его середину и на службу куда более важной задаче — созданию первой самореплицирующейся молекулы РНК: рибозимы-полимеразы гораздо эффективнее копируют зеркальные варианты друг друга, чем свои собственные! Вряд ли можно представить более наглядную иллюстрацию этого, лежащего в самой основе жизни процесса, чем знаменитая литограмма М.К.Эшера «Рисующие руки», изображенная в самом начале статьи, где Художник совершенно верно изобразил разные руки: видно, что для нижней руки, развернутой кнаружи, этот процесс затруднен — а значит она не сможет нарушать процесс копирования верхней, поскольку одна и та же молекула не может быть одновременно и матрицей и копирующей машиной. Что ещё интересно в вышеуказанной статье о рибозмах-репликазах: длина цепи нужной молекулы РНК оказывается более чем в два раза короче, чем в случае всех предыдущих подходов — около 83 нуклеотидов (практически как у вышепоказанной транспортной РНК), тогда как в «Логике Случая» в качестве ориентировочного максимального предела «неуменьшаемой (нередуцируемой) сложности», который может быть собран в нашей Вселенной просто случайным перебором нуклеотидов с вероятностью, не превышающей количество имеющихся ресурсов и времени Е.Куниным была установлена длина уникальной последовательности в 102 нуклеотида, причем в случае рассматриваемой работы указано, что полученный там — не единственный и даже не самый эффективный вариант работающей последовательности! Более того: околотопологические подходы к проблемам хиральности, узлов, фракталов и пр. в применении к задаче построения самокопирующихся молекулярных машин вообще оказываются неожиданно продуктивными. Таким образом сложная химическая жизнь возможна только на базе нерегулярных хиральных гетерополимеров. Ясно, что сложные машины из конструктора с правильным набором деталей подручными средствами или даже путем случайных перемешиваний сделать гораздо проще, чем из большого количества простых, даже высококачественных, но никак не обработанных материалов. Известная метафора креационистов о невозможности урагана на свалке собрать из мусора Боинг-747 неадекватна еще и по этой причине. Чтобы приблизить её к реальности лучше говорить о торнадо, переворачивающем склад SNOT-элементов фабрики LEGO, причем на регулярной основе — а в этом случае результат уже не так однозначен. Особенно если эти SNOT-блоки снабдить сильными магнитами. И в определенном смысле роль таких «магнитов» — участников и посредников полярных взаимодействий в организмах выполняют ионы. В основном — ионы металлов. Причем не следует думать, что все они — микроэлементы. Прежде всего это касается калия. А вот более распространенный в океанах (причем не только на Земле) натрий — содержится внутри клетки в куда меньших количествах (рис. 4), что является одним из нагляднейших свидетельств в пользу зарождения жизни не в океане, а в геотермальных полях. Однако в следующей своей статье те же авторы совершенно ясно показали первичность натриевой энергетики перед протонной (жизнь нередко довольно «диалектично» устроена). Рис.4. Эссенциальные (необходимые) элементы, используемые земной биотой. 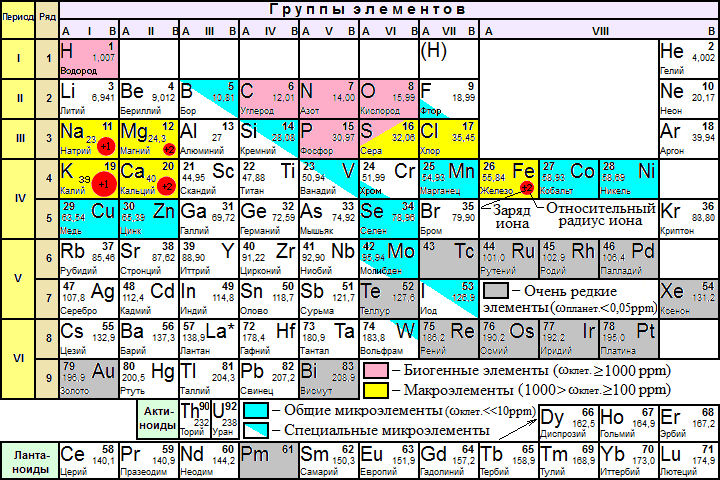 Как видно из таблицы, известная нам жизнь уже задействовала целую треть всех стабильных и более-менее распространенных элементов (24 из 72): практически все сколь-нибудь широкораспространенные неметаллы, и ряд металлов вплоть до весьма редкого молибдена, без которого эффективный механизм фиксации вышеупомянутого азота из атмосферы для постройки своего тела эволюция так и не смогла создать (более распространенный ванадий или даже просто железо тоже позволяют фиксировать азот, но гораздо хуже). Другой часто описываемый вариант альтернативной биохимии — замена биорастворителя — воды. Сейчас некоторые исследователи возлагают определенные надежды на холодные планеты с углеводородными океанами, как на Титане, что встречает очень серьезное препятствие: углеводороды — практически неполярные молекулы — то есть электронная плотность в них распределена довольно равномерно: «более отрицательно» или «более положительно» заряженных мест в них не выделяется. Неполярные же молекулы хорошо растворяются только в таких же неполярных растворителях, но практически нерастворимы в полярных (и наоборот). И на этих взаимодействиях и с учетом их построены все структуры в клетках — на этом сочетании базируется почти вся молекулярная самоорганизация. И, хотя химическое моделирование допускает существование инвертированных клеточных мембран, например, из акрилонитрила — этот факт доказывает только то, что найти основу для мембраны — далеко не самая сложная задача для образующейся жизни. Хотя само наличие гибкой полупроницаемой оболочки, все же необходимо для появления любых хоть сколь-нибудь сложных организмов (сложнее вирусов), иначе мы снова встаём перед проблемой нередуцируемой сложности как в случае с кристаллами, когда жесткая оболочка потребует сразу слишком большое число транспортных систем для обмена всеми нужными разновидностями веществ со внешней средой. Причем те автономные организмы, которые некогда пошли по пути образования малопроницаемых оболочек, хотя и существенно повышают устойчивость во внешней среде, в то же время на несколько порядков теряют в скорости роста и размножения и в результате имеют довольно примитивное строение. Наглядный пример: наши микобактерии (в их числе — возбудители туберкулеза и лепры). Главная проблема неполярных сред в том, что практически любая органическая молекула с несимметрично расположенным гетероатомом (кислорода, азота, фосфора или любого другого элемента кроме углерода и водорода) будет заметно полярнее растворителя. И сами по себе полярные взаимодействия — намного более необходимы для обеспечения разнообразия чем даже упомянутые в начале главы кратные пи-связи. И, поскольку сила связи полярных молекул существенно больше чем неполярных, то при температуре жидкой метан-этановой смеси любые слегка полярные органические молекулы крупнее акрилнитрила будут выпадать на дно в виде нерастворимых кристаллов. Причем этот эффект давно и широко применяется в виде фракционирования углеводородов и спиртового осаждения нуклеиновых кислот (ДНК не растворяется даже в довольно полярном этиловом спирте!). Теперь, сконцентрировавшись на полярных растворителях, первое на что обращается внимание — это аммиак. Этот вариант ограничен не только и не столько более узкой «зоной жизни» для гипотетических планет с аммиачными океанами вследствие того, что жидкий аммиак (NH3) существует при меньшем диапазоне температур (при стандартном давлении — вдвое по сравнению с водой), и даже не тем фактом, что ультрафиолетовому расщеплению (фотолизу) NH3 подвергается гораздо легче чем H2O — если планета достаточно крупная (суперземля), то водород не будет быстро улетучиваться из атмосферы и процесс станет обратимым. Главная проблема аммиака как растворителя в том, что образующий его азот — менее электроотрицательный и при этом на порядок менее распространенный элемент по сравнению с кислородом, а значит он будет проигрывать последнему конкуренцию за нещелочные металлы, которые присутствуют на планетах главным образом в виде практически нерастворимых в нём оксидов и комплексных солей оксикислот — как кислота аммиак в десять порядков слабее воды. Это не говоря о том, что само существование жидкого океана аммиака без существенных примесей воды вообще довольно маловероятно по причине того, что физические свойства этих веществ очень сходны. В то же время достаточно концентрированный раствор аммиака в воде не только может быть, но скорее всего и был исходной средой для биогенеза известной нам жизни: дело в том, что размер ионов калия K+ — самого распространенного металла в клетках (доли калия и серы там практически равны) очень близко совпадает с размером ионов аммония NH4+, образующихся при растворении аммиака в воде (особенно подкисленной). А сортировку ионов одного заряда через мембрану клетки могут осуществлять только по их размеру. Таким образом транспортные системы клеток практически не различают ионы калия и аммония. Более того: было показано, что ионы аммония способны не только полностью заменять ионы калия в бактериальных клетках (при сохранении кислотно-основного баланса — pH) без потери их жизнеспособности, но и показывают бОльшую эффективность в синтезе белка! А тот факт, что эвтектические смеси аммиака с водой имеют существенно меньшие температуры плавления — вплоть до 175K (-100°C) при соотношении 2:1 ("дигидрат аммония") ещё и заметно расширяет зону жизни. Недавно был предложен ещё один вариант альтернативного биорастворителя — сверхкритический или жидкий диоксид углерода: показано, что даже в безводном сверхкритическом CO2 белки-ферменты сохраняют свою каталитическую активность. Причём дольше, чем в водном растворе той же температуры. С другой стороны — их части становились менее подвижными. Поэтому такая среда кажется подходящим биорастоворителем для высокотемпературных сред. Это кажется довольно странным: ведь CO2 — неполярная молекула. Однако она способна в некоторых пределах поляризоваться под высоким давлением. Но даже в этом случае scCO2 способен растворять только довольно малополярные молекулы и для повышения растворимости полярной органики в него добавляют модификаторы — обычно простые спирты — метиловый или этиловый. На первый взгляд и это не такой уж критичный момент — многие вспомнят неоднократно мелькавшие новости про обнаружение «гигантских облаков спирта в космосе», и действительно — указанные соединения в межзвездном пространстве не редки. Но не известно — способны ли они пережить образование планет. И снова, как и в случае с аммиаком, можно вспомнить о ещё более вездесущей и стабильной воде. И здесь мы также видим замечательное сродство этих веществ: углекислый газ весьма неплохо растворяется в воде, где он, вступая с ней в обратимую реакцию образует очень даже полярную молекулу угольной кислоты. А её ион — гидрокарбонат HСO3- — самый распространенный отрицательно заряженный ион (анион) в известных нам живых клетках. А вовсе не хлорид-ион Cl-, избыток которого, например, как знает всякий агроном, не менее вреден чем и избыток ионов натрия Na+ для тех организмов, которые не могут сами сменить среду обитания — прежде всего для растений (засаливание почвы). Более того — если рассмотреть роль хлора в одноклеточных организмах (особенно прокариот), то ничего специфически важного и незаменимого кроме компенсации pH в присутствии сильных катионов щелочных металлов — калия и натрия (с чем не может справиться в одиночку более слабый гидрокарбонат-ион) он не делает! Теперь же, если мы вернемся к иону аммония, то увидим, что он тоже образует гораздо более слабое основание чем ион калия: pH насыщенного раствора «гидроксида аммония» никогда не превышает 12 единиц, которые способны переживать (хотя и с трудом) современные экстремальные алкалофильные прокариоты-рекордсмены. Что полностью снимает необходимость в компенсации pH сильным хлорид-анионом — достаточно, чтобы над раствором было достаточно углекислого газа, из которого почти только и состоят атмосферы всех известных нам землеподобных планет — Венеры, Марса и, конечно, самой древней Земли. (Причем, что интересно: их соединение — соль гидрокарбонат аммония в сухом виде достаточно стабильное твердое вещество, распадающееся при нагревании до... 42°C — при этой температуре также начинают денатурировать некоторые белки высших организмов.) Завершая тему растворителей, следует отметить ещё одну важную вещь: все три рассмотренных вещества: аммиак, диоксид углерода и вода для многих современных организмов являются единственными и довольно легкоусвояемыми источниками азота, углерода и кислорода соответственно для построения всей своей органики. Также все три вещества являются полярными или поляризуемыми растворителями, жидкими в сходном диапазоне температур и очень широко распространенными во Вселенной. Поэтому, принимая во внимание ранее описанный «модульный принцип» построения биополимеров следует заключить: нет никакой «водной», «аммиачной», «углекислотной» жизни, потому что для возникновения очень сложной органики из которой только и может появиться жизнь, нужно присутствие всех трех указанных веществ в сравнимых количествах или их производных — таких как формамид и цианиды. Коснемся теперь такого процесса как катаболизм — т.е. окисление и восстановление различных веществ с целью извлечения энергии. Использование в качестве окислителя (акцептора электронов) молекулярного кислорода (O2) всей сложной (эукариотической) жизнью также не случайно: по все той же причине его исключительно высокой электроотрицательности, соединения, содержащие в своем составе соседей по таблице элементов — азот и серу — обладают более низким окислительным потенциалом. Хотя и этот вариант обмена веществ земной жизнью тоже очень давно освоен и активно используется в бескислородных средах. Причем сера является там как продуктом бескислородного фотосинтеза, так и окислителем для анаэробного дыхания. Оксиды азота тоже издревле используются бактериями и как акцептор и как донор электронов (а высшими организмами — и как сигнальная молекула). Но его широкому распространению, кроме меньшей эффективности по сравнению с кислородом, вероятно, помешал ещё и тот факт, что азот очень трудно добыть из его атмосферного простого вещества (N2), а прочие источники его на нашей планете давным-давно были фактически исчерпаны (последние ускользнувшие от биосферы ископаемые остатки селитры были добыты человеком в первой половине прошлого века). Поэтому практически весь фиксируемый организмами азот идет на синтез биополимеров, составляющих их тела. Подводя итог главе, следует заключить: появлению альтернативной биохимии препятствуют два фактора: редкость элемента, либо неэффективность его в качестве замены (а часто — и то и другое сразу): с кислородом в качестве окислителя может конкурировать только фтор, но для этого его слишком мало как на нашей планете, так и во всей известной вселенной в целом, хотя достаточно для минеральной основы зубной эмали. Хлор же, не смотря на то, что он гораздо более распространен и составляет почти 1/50 массы морской воды, имеет меньшую электроотрицательность по сравнению с кислородом и, вероятно, не очень удобен для биохимии из-за способности образовывать всего одну стабильную связь. Тем не менее, некоторые морские организмы оказались способны синтезировать разнообразные хлорорганические соединения и другие органогалогены, ранее считавшиеся «ксенобиотиками» — веществами, совершенно чуждыми биосфере и опасными для большинства организмов. Приблизительно ту же картину мы видим и в случае второго (после кислорода) по распространенности элемента земной коры — кремния: не смотря на чрезвычайную распространенность, живые организмы не спешат включать его в свои тела: в небольших количествах его, наряду с бором, используют растения для образования межклеточных связей, строят панцирь одноклеточные эукариоты: диатомовые водоросли и радиолярии, и образуют скелет некоторые губки. Т.е. жизнь не только была способна использовать большинство доступных элементов когда-либо за свою историю, но и успешно демонстрирует эту способность по сей день, однако — в очень ограниченном масштабе. Этот факт даже самому упрямому энтузиасту биохимической экзотики должен наглядно показать справедливость поговорки: не все то золото, что в одной с ним подгруппе. 3. Планетология и астрономия биологии (пространственные ограничения) "Строительные материалы жизни", о которых шла речь в предыдущем разделе, чтобы обеспечить её распространение и, тем самым, — длительное существование и развитие — должны быть доступны в планетарном масштабе. И такую доступность, с учетом того, что жизнь зарождается не в излишне соленом океане, а в теплых прудах Дарвина — геотермальных источниках, можно обеспечить только через атмосферу (которая также служит и дополнительным, практически неисчерпаемым буфером для реакционной среды) — т.е. в газообразной форме, каковую на нашей планете прекрасно демонстрируют и поныне все три вышеуказанных очень полярных материала-растворителя, а вторую половину её истории — ещё и сильнейший из доступных окислитель (молекулярный кислород). Отдав должное гидросфере и атмосфере, ни в коем случае нельзя забыть и о роли литосферы: синтез ключевых органических соединений-предшественников биомолекул не мог происходить без целого ряда разнообразных катализаторов, среди которых подавляющее большинство — и тогда и сейчас, и в живых клетках и в промышленном синтезе — составляют многовалентные металлы: железо, марганец, цинк, кобальт и др. Эти металлы на заре жизни работали не только и столько в виде растворённых ионов (большинство их солей вообще малорастворимы), сколько в виде твердых пористых пород двухвалентных оксидов и сульфидов, которые стали, по сути, минеральной матрицей для фиксации и сборки как молекул-предшественников, так и целых биополимеров. Эти конкретно-химические аргументы в пользу зарождения жизни в дарвиновском «теплом пруду» на суше и против океанической гипотезы подкрепляются еще и вескими более общими соображениями: 1) В большом водоеме происходит слишком сильное разбавление молекул, чтобы они могли эффективно взаимодействовать друг с другом. 2) Единственная молекула-саморепликатор может быть очень эффективна, но из-за полной самодостаточности будет иметь и очень малый потенциал для эволюции, а комплексы-гиперциклы взаимодополняющих молекул уязвимы к эгоистам-паразитам, от чего можно избавиться путем пространственной изоляции разных наборов молекул в «микрореакторы», которыми могут служить поры минеральных пород (запустив тем самым групповой отбор). 3) Чем меньше объем «реакционных камер» — тем больше площадь соприкосновения катализатора с реакционной смесью. Поэтому зарождение жизни в океане если и возможно, то гораздо скорее это произойдет в приповерхностной толще донных пород. Причем пристрастие её к границам раздела фаз замечательно прослеживается и поныне: от бактерий, демонстрирующих свое самое сложное поведение в виде колоний на плотной среде, а также в виде биопленок, которые являются самой распространенной формой их существования «в дикой природе» (а вовсе не взвеси планктонных клеток!) и настоящих бактериальных матов — первого полностью достоверного наиболее древнего свидетельства жизни (и скорее всего — самой массовой её формы в ту эру) до планетарного распределения биомассы, океанская часть которой не только по суммарному количеству, но и по видовому разнообразию меньше чем на суше, причем почти вся океанская биота обитает в узкой прибрежной и приполярной зоне на мелководье. Раз уж в возникновении жизни приняли непосредственное участие «вода», «воздух» и «земля», то должно бы найтись место и «огню» в соответствии с учением Парацельса о «квинтэссенции», где наблюдатель возникает как «вытяжка» сложности всех первоэлементов целого мира. И действительно: «огонь» в форме ультрафиолетового излучения Солнца вполне годится на место как одного из ключевых источников химического разнообразия, так и фактора отбора азотистых оснований для наших нуклеотидов. Таким образом, хотя зарождение жизни и происходит только в жидкой полярной многокомпонентной среде, но только в теснейшем взаимодействии с газообразной атмосферой и твердыми минеральными матрицами. Что же касается требований к основному источнику энергии на планете — центральному светилу системы (или нескольким светилам — если звезд в ней несколько), то оно хорошо известно: это стабильность на протяжении хотя бы нескольких миллиардов лет. У самых долгоживущих звезд — красных карликов — расположение зоны жизни (области жидкого состояния полярных растворителей) находится внутри области приливной синхронизации вращения, и всякая пригодная для жизни планета будет всегда повернута к своему светилу одной стороной — подобно Меркурию к Солнцу и Луне к Земле. В то же время модели показывают, что планеты с достаточной плотностью атмосферы, а также объемом и площадью гидросферы, вполне способны обеспечить выравнивание температур на освещенной и теневой сторонах для существования жидких полярных растворителей. Способность удержать атмосферу особенно важна с учетом мощных и частых вспышек этих звезд. Поэтому наиболее подходящим типом планеты для жизни в такой системе должны стать «суперземли», большинство из которых, вероятно, целиком покрыты глубоким океаном. Но сильно глубокий океан наработке биомассы и биоразнообразия не способствует, как нам показывает пример нашей собственной планеты. Либо же на дне должна существовать весьма бурная тектоническая активность литосферы для обеспечения хемосинтетических биоценозов, аналогичных нашим «черным курильщикам». Если же океан будет слишком глубоким — более сотни километров — кора может быть практически полностью заблокирована экзотическим льдом VII и VI высокого давления, что сделает полностью невозможным ни существование хемосинтетиков, ни само зарождение жизни в таких условиях. Но даже если планета будет иметь сушу, выход на нее жизни возможен только в области терминатора, с более-менее комфортными температурными условиями, но при этом — постоянными и очень сильными ветрами, что не позволит развиться ничему крупнее лишайников. Кроме того, взятие проб с такой планеты станет очень непростым делом: спускаемый аппарат должен сразу приводняться и потом стартовать с воды же. В то же время для спутников планет-гигантов, вращающихся в этой зоне, проблема синхронизации вращения не существует, что может сделать их значительно более подходящими кандидатами для экзобиологических исследований при соблюдении прочих условий. Поэтому поиск жизнепригодных планет в обозримой перспективе имеет смысл ограничить только карликами Главной последовательности (классов G, K, M), причем расположенных не ближе нескольких десятков световых лет от остатка сверхновых (в зависимости от мощности взрыва) — нейтронных звезд или черных дыр. Кроме подробного разбора вопросов планетарного возникновения жизни, необходимо также сказать пару слов и о гипотезе внепланетного возникновения жизни — панспермии. Хотя сегодня уже довольно трудно представить себе условия, которые бы благоприятстовали возникновению бурной биохимии в условиях открытого космоса, ряд недавних работ все еще не позволяет окончательно отвергнуть предположения о том, что хотя бы некоторые этапы ранней эволюции протекали во внепланетарных условиях. Что же касается переноса уже вполне сформировавшейся жизни между телами в пределах одной звездной системы, то проведенное моделирование показало весьма высокую вероятность данного сценария. Однако эта модель не отвергает возможность зарождения жизни на планете, а только ещё более осложняет задачу поиска источника жизни, попросту разнося её по всей звездной системе: так, например, мы не можем сейчас полностью и окончательно исключить возможность зарождения нашей жизни где-нибудь на древнем Марсе или даже на Венере (если на ней вообще когда-то существовала вода, что сомнительно) и уже вторичный её заброс на Землю (хотя это все же значительно менее вероятно, чем земной геотермальный сценарий). В то же время, высокая вероятность «рассева» жизни по звездной системе автоматически подразумевает заселение всех крупных объектов в пределах зоны обитаемости, и отсутствие каких бы то ни было признаков жизни на Марсе показывает, что потенциал адаптации у неё хотя и весьма широк, но отнюдь не безграничен. 4. Биология² (временные ограничения) Если доступные нам методы всё ещё не позволяют полностью рассеять тьму неопределенности химических основ живых организмов, то на что же мы должны рассчитывать при поиске «под фонарем» — у структурно близких аналогов жизни земного типа? Следует отметить: хотя земная жизнь и совершенно ничтожна в пространстве Вселенной, она существует уже целую треть времени, прошедшего от момента Большого Взрыва! (Поэтому поэтические строки, вынесенные в эпиграф — совершенно точны с космологической точки зрения.) А теперь давайте повнимательнее рассмотрим временную шкалу с этого момента: Прежде всего интересен тот факт, что известная нам жизнь в виде уже практически современных прокариотических (бактериальных и архейных) сообществ обнаруживается на Земле очень скоро после её образования (более 4,1 млрд. лет назад), а по факту — в самых древних доступных осадочных (с достоверными следами воды) породах (более 3,8 млрд. лет назад) — т.е. весь абиогенез, если он происходил здесь, — полностью завершился не более чем за полмиллиарда лет. Но не следует думать, будто прокариотическая клетка — самодостаточная форма жизни: ни одна линия организмов не сможет существовать миллионы и миллиарды лет, если не будет достаточно часто обмениваться генами с другими линиями. У прокариот же полового размножения нет, а половой процесс (коньюгация), как отмечает Евгений Кунин в «Логике случая»(с.160), известен только у очень ограниченного числа бактерий и вовсе не обнаружен у архей. Другой способ переноса генов — поглощение свободноплавающего генетического материала из окружающей среды (трансформация), не смотря на чрезвычайно широкое применение в биоинженерии, даже в лаборатории происходит с невысокой вероятностью: даже в идеальных и очень кратковременных условиях трансформируется только одна клетка из нескольких тысяч (этот известный факт проверил собственноручно даже автор этих строк). В итоге остается только один способ генетической рекомбинации — «принудительный» — при помощи вирусных частиц. В свете того, что вирусы бактерий (бактериофаги) оказались самыми многочисленными и чрезвычайно разнообразными формами жизни в океане, это тем более не удивительно. Таким образом, не только вирусы не могут без клеток, но и простые, прокариотические клетки не способны не только развиваться, но и просто сохраниться в конкурентной среде достаточно длительный срок без обеспечиваемого вирусами горизонтального переноса генов. В целом можно даже сказать, что вирусы бактерий, биологическая роль которых — распространение генов, являются аналогом... самцов многоклеточных эукариот. Вирусы возникли одновременно с клетками, сопровождая их становление и развитие всю историю живого мира, особенно на самом раннем, протоклеточном этапе, когда «полуклетки» были неразрывно связаны с минеральной матрицей-катализатором и тем более были неспособны к самостоятельному обмену генами, когда он был особенно важен. На счету вирусов — длинный ряд важнейших инноваций — от самой ДНК до плаценты высших млекопитающих. Вместе с тем, вирусы всегда составляли отдельный «мир»: гены вирусного происхождения как правило очень хорошо отличимы от клеточных генов. То же касается и их вирусных продуктов и прежде всего — белков оболочки (капсида). В отличие от клеточной мембраны, оболочка вирусной частицы (вириона) представляет собой совершенно жесткий каркас, состоящий только из белка и играющий роль конверта или посылки для наследственного материала, выполняя только три функции: сохранение содержимого ("упаковка"), распознавание и прилипание(адгезия) к подходящей клетке ("адрес на упаковке") и внедрение содержимого ("лезвие в комплекте"). Таким образом, если мы снова обратимся к рассуждениям из главы про химию о требованиях к оболочке простейшего организма, то снова увидим — как в случае с аммиаком, углекислотой и кристаллическими матрицами — что природа и здесь тоже уже задействовала и такое решение как жесткая оболочка — настолько, насколько это вообще может быть применимо в живой системе естественного происхождения. Совокупность этих фактов подводит нас к мысли о том, что «архитектурная» основа земной жизни — не частный случай вселенского многообразия, но одно из очень и очень немногих решений комбинаторной головоломки космического масштаба. Следующий очень важный этап развития жизни: появление эукариот — клеток современного типа с куда более сложной организацией: 1) С разделением внутриклеточного пространства на изолированные камеры с помощью мембран — таких же как поверхностная, но не сязанных с ней ("компартментализация"), среди которых самая сложная и большая — это ядро, всегда содержащее несколько очень плотно и сложно упакованных молекул ДНК — хромосом. 2) Очень развитым белковым цитоскелетом, связывающим цитоплазму настолько, что она даже не вытекает при небольшом проколе мембраны (т.е. это уже по сути не раствор, а гель, пронизанный множеством молекулярных транспортных систем). 3) Содержащих в себе остатки симбиотических бактерий — митохондрии, которые обеспечивают в 18 раз более эффективное получение энергии за счет молекулярного кислорода, что мы называем дыханием. 4) И просто на два-три порядка более крупным размером. Все четыре признака по отдельности, конечно, можно встретить и у отдельных прокариот: - внутриклеточные мембранные тилакоиды цианобактерий или несколько линейных молекул хромосомной ДНК у бореллий; - цитоскелет под мембраной у всех палочковидных и спиралевидных бактерий; - тесный симбиоз (в форме паразитизма) у наноархей и бделловибрионов на других, более крупных археях и бактериях соответственно; - огромные размеры у морской бактерии тиомаргариты. Но в несравненно более простом виде, тогда как все известные эукариоты имеют все перечисленные признаки в совершенно равной и полноценной форме (за исключением разве что глубокопаразитических примитивнейших грибов-микроспоридий, у которых митохондрии сильно деградировали за ненадобностью). Кстати ближайшие ныне живущие родственники самих митохондрий — это пелагибактерии (вероятно, самые распространенные в океане и на Земле прокариоты с самым маленьким геномом, позволяющим самостоятельно жить в «дикой» среде) и риккетсии (возбудители сыпного тифа). Самые древние следы эукариот (причем уже колониальных водорослей!) обнаруживаются пока не ранее 2,1 млрд. лет назад. Т.е. более чем через ~1,75 млрд. лет после первых следов на Земле жизни вообще. Это значит, что на эволюцию жизни от элементарных прокариотических клеток до клеток эукариотических (на основе которых построены все многоклеточные организмы: грибы, растения и животные) потребовалось в 3,5 раза больше времени чем на возникновение жизни вообще — если мы считаем, что возникла она на нашей планете (от вышеуказанных 4,2 млрд. лет) или в 2,2 раза — если мы придерживаемся теории панспермии: внеземного абиогенеза с последующем заносом только что сформировавшейся клеточной жизни (от конца поздней тяжелой бомбардировки в в катархее). Такая «непрогрессивная задержка» может стать довольно досадным фактом для апологетов (сторонников) универсального самоускоряющегося прогресса, стремящихся экстраполировать (продлить) наблюдения типа «закона Мура» за область их непосредственного применения. Следующий же этап усложнения органического мира — возникновение многоклеточности — датируется всего 0,85–0,64 млрд. лет назад. Т.е. не позже чем через 1,2 млрд. лет после эукариотизации. Причем, в отличие от последней, у растений она произошла независимо от животных и далеко неоднократно. И здесь линия ускоряющегося прогресса как будто бы начинает выравниваться. Это положение вещей будет казаться куда менее странным, если обратиться к различию взгляда на сложность живых организмов и отдельных изобретений эволюции на основе опыта специалиста и интуиции неспециалиста, сведенные в такую таблицу: Сравнительная сложность биологических механизмов
Индекс интуитивной сложности вычислен на основании опроса четырех десятков неспециалистов, где за единицу взято число голосов за наименее популярный вариант, которым оказались ложноножки амебы. Т.е. самым сложным из представленных механизмов считают «жгутик» в три раза больше опрошенных, чем число голосовавших за «ложноножку». В то время как реальная сложность механизма может быть оценена только числом уникальных деталей из которого она состоит, что прямо отражено в числе генов, эти детали (в данном случае — конкретные структурные или регуляторные белки) кодирующих. Как видно: любимый креационистами «уникальный» якобы «нередуцируемо-сложный» бактериальный жгутик — механизм того же порядка сложности, что и «хаотично-аморфная низкоорганизованная» ложноножка амеб — на самом деле одних из наиболее сложно устроенных одноклеточных эукариотических организмов (после инфузорий) и имеющих одни из самых больших геномов среди всех организмов вообще (включая многих позвоночных и цветковые растения!) Помимо школьной программы биологии (где до недавнего времени прокариоты (и бактерии в частности) не упоминались вообще, а изучение зоологии начиналось сразу с «простейшей» (в буквально-классификационном, но отнюдь не в прямом смысле!) ни в чем не повинной амебы) в такой нестыковке виновато и наше интуитивное восприятие, сформированное закономерностями макромира — мира классической механики, где из фундаментальных взаимодействий взгляду видна почти только одна гравитация. Но совсем не так обстоит дело уже в мире насекомых, где очень важное значение начинает играть сила поверхностного натяжения (нагляднейший пример — водомерки). И тем более далек от нашей интуиции мир молекулярных взаимодействий — чрезвыайно сильных в микромасштабе. В мире таких взаимодействий куда вероятнее ожидать самосборки практически идеальной икосаэдрической (20-гранной) оболочки из одинаковых белковых молекул вокруг вирусной НК, чем выпячивания в нужном направлении «аморфной», а в действительности — состоящей из координированной работы сотен тысяч молекулярных машин, огромной по бактериальным меркам ложноножки амебы! И не исключено, что именно благодаря этой системе стал возможен активный захват (фагоцитоз) архейным предком эукариот других бактерий с последующим слиянием в единую неразрывную систему, а также и настоящее половое размножение. К счастью, кроме вышеприведенного фильма «Внутренняя Жизнь Клетки» теперь есть еще и недорогие конструкторы из сильных магнитов на основе сплавов редкоземельных металлов, которые позволяют наглядно преодолеть это непонимание. Кроме того, невозможность более раннего появления эукариот можно объяснить не только сложностью изменений, но и необходимостью накопления достаточного количества сильного окислителя в окружающей среде, чтобы они стали выгодны. Тем более, что как теперь известно — это был довольно длительный процесс, который начался не позднее 2,4 млрд. лет назад. Также нужно упомянуть и о необходимости обеспечиваемого железным ядром хорошего магнитного поля, защищающего от основной части наиболее жесткого излучения. Так или иначе, но теперь должно быть гораздо понятнее: почему для современных биологов проблема возникновения эукариотической клетки стоит сразу за проблемой возникновения жизни, а многоклеточность и отдельные приспособления: жгутики, фотосинтез, дыхание и пр. — проблемой как таковой являются теперь в куда меньшей степени. Дальнейшая же эволюция по пути многоклеточности, как уже показано, была гораздо более простым и понятным этапом, причем происходившим неоднократно. Более того: генетический аппарат предшественников настоящих многоклеточных животных (эуметазоев) имел более чем достаточно задатков для дальнейшего увеличения сложности, причем даже у ближайших наших одноклеточных родственников (да-да: именно заднежгутиковые сперматозоиды — это то, что отличает нас от растений, водорослей, амеб, инфузорий и всяких плазмодиев и роднит с грибами!) Что-то же более-менее похожее на двусторонне-симметричных животных появляется около 0,6 млрд. лет назад. А практически все основные современные типы животных — довольно резко проявляются 0,54 млрд. лет назад ("Кембрийский взрыв"). Кроме того теперь установлено, что и нервная система возникала неоднократно. 5. Теология биологии (вместо заключения) В области вопроса о происхождении жизни за последние десятилетия были достигнуты действительно большие успехи. И тот факт, что число возможных вариантов значительно сузилось и число потенциальных форм жизни стало значительно беднее — есть прямое следствие этих успехов: точно также как великие географические открытия убрали с карты мира русалок, псоглавцев и разнообразную морскую демонологию — так и теперь уже не только ближайшие планеты оказались необитаемыми, но и концепция экзотических обитателей более далеких газовых гигантов оказалась под большим вопросом, и парадокс Ферми становится в свете этих новых знаний все менее парадоксальным. Чем больше происходит погружение в проблему происхождения жизни, тем все более она предстает не как конструктор, из которого можно собрать все, что пожелаешь и потом обозвать коллег с «недостатком фантазии» «углеродными шовинистами»; а как головоломка с очень небольшим числом правильных решений. Уже простое перечисления таких фактов об известной нам жизни, как: - использование целой трети доступных химических элементов для построения себя; - освоение третьей части температурной шкалы от абсолютного нуля; - существование трети времени от начала вселенной; - освоение не только полужидких, но и твердых кристаллоподобных структур вирусных капсидов, микротрубочек и белковых волокон; должно прямо наводить на мысль, что жизнь вообще — не самое заурядное явление во Вселенной. Это было понятно даже алхимику Парацельсу, ибо сложная система может появиться (и развиваться!) только в исходно сложной среде, а система самой высокой сложности из известных нам — сам наблюдатель — должна быть, по сути, квинтэссенцией сложности Мира. И называть её «водно-углеродной» — это совершенно некорректное опрощение. Поэтому на смену некорректному и давно устаревшему термину «водно-углеродного шовинизма» предлагается более широкое и одновременно более точное понятие «полярно-комбинаторного шовинизма», которое теперь, подобно «центральной догме молекулярной биологии», должно быть свободно от негативных коннотаций старых концепций биогенеза. В то же время тот факт, что жизнь нам известна только в одной форме, продолжает давление на методологию биологии как естественной науки, которая, чтобы не стать разделом истории (т.е. пустым описательством фактов), в нормальном случае должна иметь дело только с повторяемыми явлениями. Если внутри самой биологии небольшие и средние усложнения (те же многоклеточность и нервная система) могут возникать несколько раз параллельно, то по поводу наиболее крупных (эукариотизации и самого зарождения жизни) у нас всё ещё очень мало твердых доказательств их исторической неуникальности. И если бы можно было запустить эволюцию заново даже практически в тех же условиях — мы совсем не можем быть уверены, что и крупные таксоны (типы и классы) были бы такими же как мы их знаем или были бы вообще. Так «вопрос ребром» был поставлен в книге «Логика Случая» Евгения Кунина, но, хотя в самое последнее время уже появилось немало прекрасных работ, существенно срезающих наиболее острые углы гипотезы «невероятности жизни», — пока мы не увидим другую жизнь в пробирке или в космосе, вопрос нашей (не)уникальности так и будет висеть на прежнем месте. Но и эту печаль, даже если мы не можем разрешить ее сейчас, можно (подобно проблеме хиральности) тоже попытаться развернуть в свою пользу хотя бы как источник формальных определений для фундамента теоретической биологии. Например можно взять тот же кунинский «предел невероятности», выражаемый в такой длине какой-либо уникальной нерегулярной последовательности, вероятность возникновения которой в пригодных для этого средах и за все время их существования во всей известной Вселенной меньше единицы (для РНК это будут те самые ~102 нуклеотида). Или, к примеру, брать просто последовательность такой длины, что число ее вариантов больше числа элементарных частиц обычной (барионной) материи в той же Вселенной (для РНК это число будет немного побольше — 132 нуклеотида). Правда второй способ уже совсем условный, но в случае, когда мы не можем определить ни количество ни время существования пригодных для её появления сред — это самый простой выход. Таким образом, системы дарвиновских матричных репликаторов с уникальными последовательностями меньшей длины можно условно называть «преджизнью», а преодолевших «предел невероятности» — уже настоящими «живыми организмами». Убежден: с таким подходом вполне можно отправляться на поиски иной жизни как в глубь космоса, так и в глубь пробирки или компьютера. Андрющенко Сергей aka «Rattus the Contaminator»
2015 год
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Дизайн Elite Games V5 beta.18 EGM Elite Games Manager v5.17 02.05.2010 |
")